
Будова бактерій
Основна стаття: Структура бактеріальної клітини
[ред.] Внутрішня структура
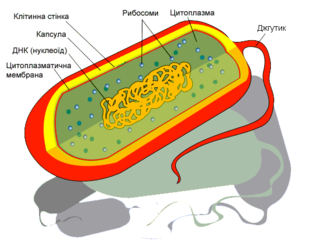
Структура клітини прокаріотів
Внутрішня частина бактерії — цитоплазма — охоплюється одною або двома мембранами, що відділяють її від зовнішнього середовища. Внутрішня з цих мембран називається цитоплазматичною мембраною. У випадку двох мембран, друга мембрана називається зовнішньою, а простір між мембранами — периплазмою. Гомогенна фракція цитоплазми, що містить набір розчинних РНК, білків, продуктів і субстратів метаболічних реакцій, називається цитозолем або гіалоплазмою. Інша частина цитоплазми представлена різними структурними елементами, що включають хромосому, рибосоми, цитоскелет та інші, характерні для окремих видів, немембранні структури, наприклад, газові везикули. Деякі бактерії формують внутрішньоклітинні гранули для зберігання живильних речовин, таких як глікоген[14], поліфосфат [15], сірка[16] або полігідроксиалканоати [17], що дають бактеріям можливість зберігати ці речовини для подальшого використання.
Однією з основних відмінностей клітини бактерій від клітини еукаріотів є відсутність ядерної мембрани і, найчастіше, відсутність взагалі мембран всередині цитоплазми. Багато важливих біохімічних реакцій, наприклад, реакції енергетичного циклу, відбуваються завдяки іонним градієнтам через мембрани, створюючи різницю потенціалів подібно до батареї. Відсутність внутрішніх мембран у бактеріях означає, що ці реакції, наприклад, перенос електрона у реакціях електронно-транспортного ланцюжка, відбуваються через цитоплазматичну мембрану, між цитоплазмою і периплазмою [18]. Проте, у деяких фотосинтезуючих бактерій існує розвинена мережа похідних від цитоплазматичної фотосинтетичних мембран. У пурпурних бактерій (наприклад, Rhodobacter) вони зберегли зв'язок з цитоплазматичною мембраною, що легко виявляється на зрізах під електронним мікроскопом, але у ціанобактерій цей зв'язок або важко виявляється, або втрачений в процесі еволюції. Відомим класом мембранних органел бактерій, які більш нагадують еукаріотичні органели, але, можливо, теж зв'язані з цитоплазматичною мембраною, є магнетосоми, присутні у магнетотактичних бактерій.
Вся генетична інформація, необхідна для життєдіяльності бактерій, міститься в одній молекулі ДНК, що має форму ковалентно замкнутого кільця (бактеріальна хромосома). ДНК в розгорненому стані має довжину біля 1 мм. Бактеріальна хромосома (звичайно в одному екземплярі, тобто всі бактерії гаплоїдні, хоча в певних умовах одна клітина може містити декілька копій своєї хромосоми) в одній точці прикріплена до цитоплазматичної мембрани та поміщається в структурі, відділеній, але фізично не відокремленій від цитоплазми, яка називається нуклеоїдом [19]. Рибосоми бактерій також дещо відрізняються від рибосом еукаріотів та архей і мають константу седіментації 70S (на відміну від 80S у еукаріотів) [20].
[ред.] Структура клітинної оболонки
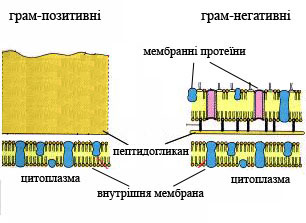
Структура клітинної стінки двох типів бактерій
Історично першою виміряною бактеріальною структурною характеристикою була структура клітинної стінки. За цією ознакою бактерії можуть бути розбиті на дві групи: грам-позитивні та грам-негативні, що визначається фарбуванням за Грамом. Грам-позитивні бактерії мають лише одну клітинну мембрану, товстий шар пептидогліканів і шар техоєвої кислоти, тоді як грам-негативні бактерії мають також зовнішню мембрану, яка містить ліпополісахариди і тонкий шар пептидогликанів, розташований в периплазмі. На поверхні, поза шаром пептидіглікану або зовнішньою мембраною, часто розташовується білковий S-шар. Цей шар забезпечує хімічний і фізичний захист поверхні клітини і може служити макромолекулярним бар'єром. S-шари мають різноманітні, але погано вивчені функції, наприклад, вони служить факторами патогенності в Campylobacter і містять зовнішні ферменти у Bacillus stearothermophilus [21].
Багато бактерій містять інші позаклітинні структури, наприклад джгутики, що використовуваються для руху клітини. Ворсинки (у тому числі фімбрії) представляють собою цілий клас незв'язаних структур, що використовуються для пересування, прикріплення до поверхонь і обміну ДНК [22][23]. Деякі бактерії також містять капсули або слизисті шари, які також полегшують прикріплення до поверхні та формування біоплівок[24]. Вони мають різну структуру від неорганізованого слизистого шару з клітинних полімерів до надзвичайно структурованих мембранних капсул. Інколи ці структури залучені до захисту клітин від поглинення клітинами еукаріотів, наприклад, макрофагами [25].
Крім того, у цитоплазматичній мембрані та клітинній оболонці розташовані спеціалізовані системи секреції, структура яких залежить від виду бактерії.
[ред.] Розмір бактерій
Бактерії можуть мати великий набір форм та розмірів (або морфологій). За розміром бактеріальні клітини звичайно в 10 разів менші, ніж клітини еукаріотів, маючи тільки 0,5—5,0 μм у своєму найбільшому розмірі, хоча гігантські бактерії, такі як Thiomargarita namibiensis та Epulopiscium fishelsoni, можуть виростати до 0,5 мм у розмірі та бути видимими неозброеним оком [2]. Найменшими вільноживучими бактеріями є мікоплазми, члени роду Mycoplasma, лише 0,3 μм у довжину, приблизно рівні за розміром найбільшим вірусам [26].
Дрібний розмір важливий для бактерій, тому що він приводить до великого співвідношення об'єму до площі поверхні, що пришвидшує транспорт поживних речовин і виділення відходів. Низьке співвідношеннях об'єму до площі навпаки обмежує швидкість метаболізму мікробу. Причина для існування великих клітин невідома, хоча здається, що великий об'єм використовується перш за все для зберігання додаткових поживних речовин. Однак, існує і найменший розмір вільноживучої бактерії. Згідно з теоретичними підрахунками, сферична клітина діаметром менше 0,15—0,20 мкм стає нездатною до самостійного відтворення, оскільки в ній фізично не поміщаються всі необхідні біополімери і структури в достатній кількості. Нещодавно були описані нанобактерії (та схожі наноби і ультрамікробактерії), що мають розміри менші від «допустимих», хоча факт існування таких бактерій все ще залишається під сумнівом. Вони, на відміну від вірусів, здатні до самостійного зростання і розмноження, але вимагають отримання ряду поживних речовин, які вони не можуть синтезувати, з середовища або від клітини-хазяїна.
[ред.] Морфологія клітини
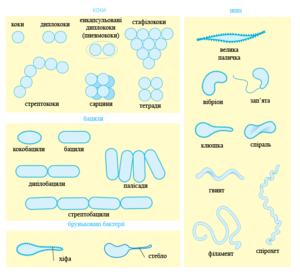
Типові форми бактеріальних клітин
Більшість бактерій мають або сферичну форму — так звані коки (від грецького слова kókkos — зерно або ягода), або паличковидну — так звані бацили (від латинського слова bacillus — паличка). Деякі паличковидні бактерії (вібріони) дещо зігнуті, а інші формують спіральні завитки (спірохети). Вся ця різноманітність форм бактерій визначається структурою їх клітинної стінки та цитоскелету [27]. Ці форми важливі для функціонування бактерій, оскільки вони можуть впливати на здатність бактерій отримувати поживні речовини, прикріплятися до поверхонь, рухатися і рятуватися від хижаків [28].
[ред.] Багатоклітинні структури бактерій
Переважна більшість видів бактерій існує у вигляді окремих клітин, тоді як інші зв'язуються разом в характерні структури, наприклад, Neisseria формує диплококи (пари), стрептококи (Streptococcus) формують ланцюжки, а стафілококи (Staphylococcus) групуються у «виноградні кетяги». Бактерії можуть також формувати нитки, наприклад Actinobacteria. Волокнисті бактерій часто оточує футляр, який містить багато індивідуальних клітин, певні види, такі як предсавники роду Nocardia, формують комплексні, розгалужені нитки, подібні до міцеліїв грибів [29].
Бактерії часто прикріплюються до поверхонь і формують щільні рогожі, що називаються біофільмами, біоплівками або мікробними плівками. Ці плівки можуть мати від декількох мікрометрів до половини міліметра в товщину і часто містять багато видів бактерій, протіст і архей. Бактерії, що живуть в біоплівках, показують складне аранжування клітинних і позаклітинних компонентів, формуючи вторинні структури, такі як мікроколонії та мережі каналів, через які постачаються поживні речовини [30][31]. У природних умовах, таких як ґрунт або поверхні рослин, більшість бактерій зв'язані з поверхнями у скліді біоплівок[32]. Біоплівки також важливі у розвитку хронічних бактеріальних інфекцій та інфекцій імплантованих медичних пристроїв, оскільки бактерії, захищені в межах цих структур, набагато складнше вбити, ніж індивідуальні бактерії [33].
Навіть складніші морфологічні зміни іноді можливі. Наприклад, при нестачі їжі міксобактерії координують свій рух, створюючи складні плодові тіла, що містять біля 100 тис. бактеріальних клітин[34]. У цих плодових тілах бактерії виконують окремі завдання, це один з прикладів найпростішої багатоклітинної організації. Частина клітин може загинути, щоб дати решті необхідні поживні речовини для закінчення процесу. Із решти частині все ж таки вдається досягти деяких частин плодових тіл, де вони діференцуються у неактивні міксоспори, стійкіші до висушування та інших несприятливих природних умов, ніж звичайні клітини, та можуть пережити голодні періоди [35].
Ще один тип багатоклітинних структур існує у деяких представників типів Cyanobacteria і Actinobacteria. У нитчастих ціанобактерій описані структури в клітинній стінці, що забезпечують контакт двох сусідніх клітин, — мікроплазмодесми. Показана можливість обміну між клітками речовиною (фарбником) і енергією (електричною складовою трансмембранного потенціалу). Деякі з нитчастих ціанобактерій містять, крім звичайних вегетативних клітин, функціонально диференційовані: акінети і гетероцисти. Останні здійснюють фіксацію азоту і інтенсивно обмінюються метаболітами з вегетативними клітинами.
[ред.] Рух
[ред.] Механізми руху у рідині
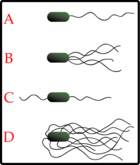
Типи розташування джгутиків:
A-монотрихи;
B-лофотрихи;
C-амфитрихи;
D-перитрихи.
Бактерії здатні пересуватися у рідині, використовуючи багато механізмів, таких як джгутики (flagella), зміни поверхневого натягу, зміни плавучості, полімеризацію актину, що оточує їх, та безліч ще невідомих механізмів.
Джгутики: Бактеріальні джгутики (flagella) — це інші структури, ніж в еукаріотів. Залежно від місця їх розташування на клітині, бактерії поділяють на монотрихи (з одним джгутиком на кінці), лофотрихи (із жмутком джгутиків на кінці) та перитрихи (з багатьма джгутиками по всій поверхні). Багато бактерій (наприклад E.coli) мають два різних режими роботи джгутиків: рух вперед та «танець» (tumbling). «Танець» дозволяє їм внести в свій рух необхідну «випадковість» [36].
Полімеризація актину: Декілька видів Listeria і Shigella рухаються усередині клітин-хазяїв, узурпуючи механізм полімерізації актину, який зазвичай використовується для руху самої еукаріотичної клітини або внутріклітинного транспорту, хоча і іншим шляхом. Просуваючи полімеризацію актину на одному полюсі своєї клітини, вони можуть формувати свого роду хвіст, який штовхає їх крізь цитоплазму клітини-хазяїна [37].
Подовжні філаменти: Деякі (спірохети) мають «подовжні філаменти», тобто джгутики, розташовані між двох мембран, та пересуваються за рахунок руху всього тіла.
Зміни плавучості: Деякі планктонні бактерії містять газові везикули, немембранні органели, що містять деяку кількость газу. За рахунок регулювання їх відносного об'єму водні бактерії можуть виконувати вертикальні міграції.
[ред.] Пересування на поверхнях
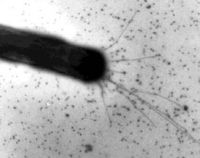
Ворсинки IV типу міксобактерії Myxococcus xanthus
Всі механізми пересування на поверхнях називаються разом бактеріальним ковзанням (bacterial gliding). Вони включають використання ворсинок та інші, ще невідомі механізми.
Ворсинки: Деякі бактерії, наприклад Myxococcus, Pseudomonas та Neisseria , рухаються за допомогою ворсинок IV типу (type four pili або TFP). При цьому методі руху ворсинки протягаються з одного полюса бактерії, зв'язуються з підкладкою та втягуються назад, створюючи значну силу у 80 пН (піконьютонів) [38].
[ред.] Регуляція руху
Рухомі бактерії можуть приваблюватися або відштовхуватися певними стимулами, така поведінка називається таксисом — наприклад, хемотаксис, фототаксис, механотаксис, магнетотаксис та еластикотаксис [39][40]. Бактерії однієї групи, міксобактерії (Myxobacteria), формують так звані плодові тіла, де вони перетворюються на міксоспори [41]. Часто бактерії використовують так зване відчуття кворуму, тобто змінюють свою поведінку залежно від щільності бактеріального населення, уповільнюючи свій ріст та інколи збільшуючи ймовірність горизонтального переносу генів.
[ред.] Ріст та генетичний апарат бактерій
[ред.] Ріст та розмноження
Див. також: Ріст бактерій
На відміну від багатоклітинних організмів, в одноклітинних організмах ріст та розмноження (поділ клітини) тісно пов'язані. Бактерії доростають до певного розміру, після чого проводять процес поділу, форму безстатевого розмноження [42]. За оптимальних умов бактерії можуть рости та ділитися надзвичайно швидко, до одного поділу кожні 9,8 хвилин для певних видів бактерій [43]. При поділі клітини створюються дві генетично ідентичні дочірні клітини. Деякі бактерії, хоча теж розмножуються безстатево, формують складніші відтворюючі структури, які полегшують поширення нових дочірніх клітин. Приклади включають створення плодових тіл міксобактеріями, створення повітряних гіф представниками роду Streptomyces та брунькування. Брунькування означає формування виступу, який пізніше відокремлюється, формуючи окрему клітину.
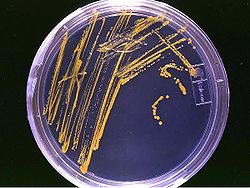
Культура бактерії на твердому середовищі (на основі агару) в чашці Петрі
У лабораторних умовах бактерії зазвичай вирощують, використовуючи тверде або рідке середовище. В якості твердого середовища використовуються чашки Петрі із шаром агару, що містить поживні речовини. Такі чашки використовуються для отримання штамів бактерій. Проте, рідке середовище зазвичай викортовується для вимірювання швидкості росту або отримання визначених об'ємів клітин. Інколи використовуються відбірні середовища, (середовища з додаванням антибіотику) для виділення та ідентифікації окремих штамів бактерій [44].
Більшість лабораторних методів зростання використовують високі рівні поживних речовин для отримання великих кількостей клітин. Проте в природних умовах кількість поживних речовин обмежена, що означає, що бактерії не можуть розмножуватися нескінченно. Це обмеження призвело до створення бактеріями різних стратегій зростання (див. теорія r/K селекції). Деякі організми можуть рости надзвичайно швидко, коли поживні речовини стають доступними, наприклад, формівання цвітіння води (за рахунок росту клітин ціанобактерій), які часто відбуваються в озерах улітку [45]. Багато організмів адаптуються до бідних та агресивних оточень, наприклад, шляхом виробництва антибіотиків представниками роду Streptomyces та іншими, тим самим не даючи рости конкуруючим мікроорганізмам [46]. Часто бактерії співпрацюють, формуючи біоплівки[32] та змінюючи швидкість росту завдяки відчуттю кворуму. Ці взаємини можуть бути істотними для зростання всієї групи організмів (синтрофія) [47].
Ріст бактерій звичайно включає три фази. Коли популяція бактерій потрапляє до багатого на поживні речовини оточення, яке дозволяє зростання, клітинам потрібний певний час, щоб пристосуватися до нового оточення. Перша фаза росту, фаза повільного росту, є фазою такого пристосування. Ця фаза характеризується високою швидкістю біосинтезу ферментів і активного транспорту [48]. За нею слідує фаза експоненціального росту, що характеризується швидким експоненціальним зростанням кількості бактерій. Швидкістю зростання вважається час подвоєння бактерій протягом цієї фази. Остання фаза росту — стаціонарна фаза, що спричинена виснажуванням поживних речовин. Клітини скорочують свою метаболічну діяльність і споживають неістотні клітинні білки. Стаціонарна фаза — це перехід від швидкого зростання до стресового стану, що характеризується збільшенням експресії генів, що беруть участь у ремонті ДНК та антиоксидантному метаболізмі [49].
[ред.] Спори та неактивні форми бактерій
Див. також: Неактивні форми бактерій
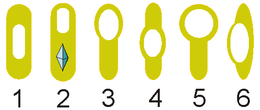
Типи морфології ендоспор. (1, 4) Центральна ендоспора, (2, 3, 5) термінальна ендоспора, (6) бічна ендоспора
Деякі бактерії типу Firmicutes здатні формувати ендоспори, що дозволяють їм витримати екстремальні екологічні і хімічні умови (наприклад, грам-позитивні Bacillus, Anaerobacter, Heliobacterium та Clostridium) [50]. Майже у всіх випадках формується одна ендоспрора, тобто це не є процесом відтворення, хоча Anaerobacter може формувати до семи ендоспор на клітину [51]. Ендоспори мають центральне ядро, складене з цитоплазми, що містить ДНК та рибосоми, оточене кірковим шаром і захищене непроникною і жорсткою оболонкою. Ендоспори не виказують ніякого метаболізму і можуть витримати екстремальний фізико-хімічний тиск, наприклад, високі рівні ультрафіолетового випромінювання, гамма-випромінювання, детергентів, дезинфікуючих засобів, нагрівання, тиску і висушування [52]. У такому неактивному стані ці організми у деяких випадках можуть залишатися життєздатними протягом мільйонів років [53][54] та виживати навіть у космічному просторі [55]. Ендоспори можуть бути причиною захворювань, наприклад, сибірська виразка може бути викликана вдиханням ендоспор Bacillus anthracis [56].
Метан-окислюючі бактерії у роду Methylosinus також формують стійкі до висушування спори, так звані екзоспори, названі так тому, що вони формуються брунькуванням на кінці клітини. Екзоспори не містять діамінопіколінової кислоти, характерного компоненту ендоспор. Іншими неактивними, оточеними товстою стінкою структури, які утворюються представниками родів Azotobacter, Bdellovibrio (бделоцисти), і Myxococcus (міксоспори) є цисти. Вони стійкі до висушування та інших шкідливих умов, але меншою мірою, ніж ендопори. При утворенні цист представниками Azotobacter поділ клітини завершується з утворенням товстої багатошарової стінки і оболонки, що оточує клітину. Нитчасті Actinobacteria формують відтворюючі спори двох категорій: кондіціоспори, які є ланцюжками спор, сформованих з міцелієподібник ниток, та спорангієспори, які формуються в спеціалізованих мішечках, спорангіях [57].
[ред.] Генетика
Більшість бактерій мають одну кругову хромосому, розмір якої може бути від лише 580 000 пар нуклеотидів у патогена людини Mycoplasma genitalium [58] до 12 200 000 пар основ в бактерії Sorangium cellulosum, що живе у ґрунті [59]. Спірохети — відомий виняток, деякі з цих бактерій, наприклад, Borrelia burgdorferi, причина хвороби Лайма, містять єдину лінійну хромосому [60]. Бактерії також можуть містити плазміди, які є маленькими позахромосомними молекулами ДНК, що інколи містять гени стійкості до антибіотиків або фактори вірулентності. Інший вид бактеріальної ДНК — інтегровані віруси (бактеріофаги). Існують багато видів бактеріофагів, деякі просто заражають і руйнують бактерію-господаря, коли інші вставляють свою ДНК в бактеріальну хромосому. Бактеріофаг може містити гени, які змінюють фенотип господаря: наприклад, при еволюції Escherichia coli O157:H7 і Clostridium botulinum гени токсину бактеріофагу перетворили безневинних спадкових бактерій на смертельних патогенних.
Mycoplasma genitalium, бактерія з найменшим відомим розміром геному
Бактерії, як безстатеві організми, успадковують ідентичні копії генів своїх батьків (тобто, вони є клонами). Проте, всі бактерії можуть еволюціонувати шляхом змін їх генетичного матеріалу (ДНК) при мутаціях. Частоти мутацій сильно залежать від виду бактерії і навіть від штаму одного виду бактерій [61]. Мутації у бактерій є наслідком як випадкових мутацій — помилок при зчитуванні протягом копіювання та впливу радіації — так і «стресових мутацій», де бактерія навмисно збільшує частоту мутацій у стресових умовах [62].
Деякі бактерії також можуть переносити генетичний матеріал між клітинами. Є три основні шляхи, як це може відбуватися. По-перше, бактерії можуть прийняти екзогенну ДНК із свого оточення у процесі, що називається трансформацією. Частіше переносяться не хромосомні гени, а плазміди. Гени можуть також бути перенесені за допомогою процесу трансдукції, коли бактеріофаг, вбудовуючись в бактерію привносить чужорідну ДНК до хромосоми. Третій метод передачі гена — бактеріальна кон'югація, коли ДНК переноситься прямим контактом між клітинами, для чого можуть використовуватися деякі типи ворсинок. Загалом всі ці шляхи переносу генетичнго матеріалу називаються горизонтальним переносом генів, і часто відбуваються за природних умов [63]. Горизонтальний перенос генів грає у бактерій роль, дещо подібну до статевого розмноження у еукаріотів. Цей процес особливо важливий у придбанні бактеріями Резистентність (стійкість) до антибіотиків, оскільки він дозволяє швидку передачу генів резистентності між бактеріальними клітинами [64].
[ред.] Метаболізм
Докладніше у статті: [[Метаболізм мікроорганізмів|Метаболізм мікроорганізмів]]

Нитки фотосинтезуючих ціанобактерій
На відміну від еукаріотів, бактерії проявляють надзвичайно широку різноманітність типів метаболізму [65]. Поширення метаболічних рис в межах груп бактерій традиційно використовувалося для визначення іх таксономії, але ці риси часто не збігаються з сучасними генетичними класифікаціями [66]. Метаболізм бактерій розрізняють за видом енергії, донора і акцептора електрона та джерела вуглецю, які використовуються для росту [67].
[ред.] Конструктивний метаболізм
За винятком деяких специфічних моментів, біохімічні шляхи, за якими здійснюється синтез білків, жирів, вуглеводів і нуклеїнових кислот у бактерій, схожі з такими ж шляхами у еукаріотів, проте за кількістю можливих шляхів і, відповідно, по ступеню залежності від надходження органічних речовин, бактерії значно переважають еукаріотів. Частина з них може синтезувати всі необхідні їм органічні молекули з неорганічних речовин (автотрофи), інші ж вимагають готових органічних сполук, які вони здатні лише трансформувати (гетеротрофи).
Найбільшим ступенем гетеротрофності відрізняються внутрішньоклітинні паразити, якщо при цьому вони здатні існувати на багатих штучних середовищах, вони називаються факультативними. Велика частина бактерій належить до сапротрофів, які незалежні від інших організмів, але використовують синтезовані ними органічні речовини. Існує також ряд бактерій, що вимагають наявності в середовищі невеликого числа певних органічних речовин (деяких аминокислот, вітамінів), які вони не можуть синтезувати самостійно (ауксотрофи), або навіть лише одного досить низькомолекулярного джерела вуглецю (цукор, спирт, кислота). Деякі з них відрізняються високою спеціалізацією (Bacillus fastidiosus може використовувати тільки сечову кислоту), інші як єдине джерело вуглецю і енергії можуть використовувати сотні різних сполук (Pseudomonas). Повними гетеротрофами з точки зору вуглецевого метаболізму можуть бути деякі фотосинтезуючі бактерії, наприклад, деякі ціанобактерії і пурпурні бактерії, які отримують вуглець з атмосферної вуглекислоти [68].
На додаток до фіксування вуглекислоти при фотосинтезі, деякі бактерії також фіксують азот із атмосфери (фіксація азоту), використовуючи фермент нітрогеназу. Ця екологічно важлива риса властива бактеріям багатьох типів [69].
[ред.] Енергетичний метаболізм
Енергетичний метаболізм бактерій засновується на фототрофії, використанні світла через фотосинтез, або на хемотрофії, використанні хімічних речовин для отримання енергії. Хемотрофи у свою чергу діляться на літотрофів, які використовують неорганічні донори електронів для дихання, і органотрофів, які використовують органічні сполуки у якості доноров електронів. Щоб використовувати хімічні сполуки як джерело енергії, електрони беруться з відновлювальних речовин і переміщуються до акцепторів електронів у процесі окислювально-відновлювальної реакції. Ця реакція вивільнює енергію, яка може використовуватися для проведення метоболічних реакцій. У аеробних організмах в якості акцептора електронів використовується кисень. У анаеробних організмах замість нього використовуються інші неорганічні речовини, наприклад нітрати, сульфати або вуглекислота. Це приводить до екологічно важливих процесів денітрифікації, відновлення сульфатів і ацетогенезу відповідно [70][71]. Ці процеси також важливі в біологічних відповідях на забруднення, наприклад, сульфат-відновлюючі бактерії в значній мірі відповідальні за виробництво надзвичайно отруйних форм ртуті (метил- і диметил-ртуть) в оточенні [72]. Анаероби використовують процес так званого анаеробного дихання (бродіння) для отримання енергії, виділяючи метаболічні побічні продукти (наприклад етанол в пивоварінні), оскільки вони не можуть бути використані далі. Факультативні анаероби можуть переключатися між процесами аеробного і анаеробного дихання, тобто різними акцепторами електронів, залежно від природних умов, в яких вони знаходяться.
Літотрофні бактерії можуть використовувати неорганічні речовини як джерело енергії. Загальними неорганічними донорами електронів є водень, чадний газ, аміак (що призводить до нітрифікації), оксид заліза II, інші відновлені іони металів та деякі відновлені сполуки сірки. Хоча це і незвичайно, деякі бактерії-метанотрофи можуть використовувати газ метан і як джерело електронів, і як субстрат для вуглецевого анаболізму [73]. Як у аеробних фототрофів, так і у хемолітотрофів, кисень використовується як кінцевий акцептор електронів, тоді як за анаеробними умовами заміть нього використовуються неорганічні сполуки. Більшість літотрофних організмів — автотрофи, тоді як органотрофи є гетеротрофами.
[ред.] Філогенетика і класифікація
[ред.] Походження
Предки сучасних бактерій були одноклітинними мікроорганізмами, які були одними з перших форм життя, що розвинулися на Землі біля 4 млрд. років тому. Протягом близько 3 млрд. років всі організми були мікроскопічними і бактерії та археї були домінуючими формами життя [74][75]. Хоча й існують бактеріальні скам'янілості, наприклад, строматоліти, відсутність у них відмітної морфології перешкоджає їх використанню для отримання даних щодо минулої історії бактеріальної еволюції або датування часуу походження специфічних видів бактерій. Проте, генетичні послідовності (у так званій молекулярній філогенетиці) можуть використовуватися для відновлення бактеріального філогенезу, і їх дані вказують, що бактерії першими відхилилися від лінії архей/еукаріотів. Останній загальних предок бактерій та архей був ймовірно термофілом, що жив 2,5—3,2 млрд. років тому [76][77], хоча можливо, що через високий рівень горизонтального переносу генів у той період слід говорити не про одного загального предка, а про популяцію.
Бактерії також прийняли участь в другому великому еволюційному розколі між археями та еукаріотами. Еукаріоти з'явилися, коли стародавні бактерії вступили в ендосимбіотичні асоціації з предками еукаріотів. Цей процес включав захоплення стародавніх альфа-протеобактерій, формуючи мітохондрій, і стародавніх ціанобактерій, формуючи хлоропласти [78][79].
[ред.] Класифікація
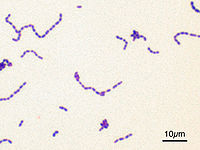
Streptococcus mutans при фарбуванні за Грамом
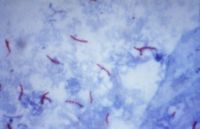
Mycobacterium tuberculosis при фарбуванні за Зіль-Нельсеном
Історично бактерії поділяли за формою на кулясті бактерії (коки, диплококи, сарцини, стрептококи), нитчасті, звивисті (спірили — форми зі спіральними завитками; вібріони, спірохети) та паличковидні. Останні об'єднювали бактерій, що не утворюють ендоспор (власне бактерії), та спороутворюючі бактерії (бацили). Але тому, що бактерії дуже дрібні та мають відносно схожі форми, класифікація, заснована на формі, не була успішною. Перша формальна класифікація з'явилася після розробки Гансом Хрістіаном Грамом методики фарбування за Грамом, що розділяє бактерії за структурними характеристиками клітинної стінки. Ця схема включає:
Gracilicutes — Грам-негативні бактерії з двома клітинними мембранами;
Firmicutes — Грам-позитивні бактерії з товстою стінкою збудованою з пептидогліканів;
Mollicutes — Грам-негативні бактерії без клітинної стінки чи другої мембрани;
Mendosicutes — Нетипово фарбовані бактерії, тепер відомо, що вони надежать до архей.
Наступний розвиток, по більшості заснований на цій схемі, включав порівняння бактерій, засновуючись на різницях їх метаболізму, виміряних за допомогою багатьох специфічних тестів. Але хоча ці схеми допомогали відрізняти штами бактерій, було неясно, чи вони відображують різницю між різними видами, чи між різними штамами того ж самого виду. Ця невпевненість була викликана відсутностю відмітних структур в більшості бактерій, також як і горизонтальним переносом генів між незв'язаними видами [80]. Більш того, саме поняття виду не може бути чітко виражене для бактерій. Ця проблема була вирішена з появою у філогенетиці генетичних методик, таких як вимірювання відносного вмісту гуаніну і цитозину, гібридизація геномів та зчитування генів (особливо гену 16S рРНК) [81]. Після цього таксономія розвилася до акуратної класифікаційної системи. У цей момент сам термін «бактерії» змінився. Термін «бактерії» традиційно застосовувався до всіх мікроскопічних прокаріотів, молекулярна систематика показала, що прокаріоти складаються з двох окремих доменів, які зараз називаються Бактерії і Археї [82]. Ці два домени, разом у Еукаріотами, стали основою системи трьох доменів, яка в даний час стала загальноприйнятою клисифікацією в біології [83]. Слід зазначити, що через деякі залишки численних історичних схем класифікації та сучасне погане знання біології бактерій їх класифікація ще не остаточна [84].
[ред.] Ідентифікація
Для початкової ідентифікації бактерій все, що широко використовується — фарбування за Грамом, яке одразу поділяє бактерії на дві великі групи та відрізняє їх від еукаріотів. Деякі організми краще всього ідентифікуються за допомогою інших методів фарбування, наприклад, кислотостійкі бактерії (Mycobacteria, Nocardia) краще всього ідентифікуються за допомогою фарбування за Зіль-Нельсеном або подібних методик [85]. Інші організми може бути потрібно ідентифікувати за допомогою вирощування їх у спеціальному середовищі, або іншими, наприклад, серологічними методами.
Методи культур дозволяють зростання тільки певного виду або групи бактерій, обмежуючи ріст інших бактерій в зразку. Часто ці методи розроблюються для специфічних зразків, наприклад, зразок слини розглядатиметься для ідентифікації організмів, які викликають пневмонію, а зразок калу розглядатиметься для ідентифікації організмів, які викликають пронос, запобігаючи зростанню нехвороботворних бактерій. Зразки, що зазвичай стерильні, наприклад, кров, сеча або спинна рідина, культивуються за умов, створених, щоб виростити всі можливі організми [86][44]. Як тільки хвороботворний організм буде ізольований, він може бути охарактеризований фарбуванням, за своєю морфологію та метаболізмом (наприклад, аероб або анаероб) та гемолітичними властивостями. Як з класифікацією бактерій, для ототожнення бактерій все більше використовуються молекулярні методи. Діагностика за допомогою таких, заснованих на ДНК, методів, наприклад, полімеразної ланцюгової реакції, набуваює популярності завдяки її специфічності і швидкості, порівняно із методами, заснованими на культурі [87].
[ред.] Взаємодія з іншими організмами
Не дивлячись на їх простоту, бактерії можуть формувати складні асоціації з іншими організмами. Ці симбіотичні асоціації можуть бути поділені на паразитизм, мутуалізм і коменсалізм. Завдяки їх дрібному розміру, бактерії-коменсали усюдисущі і ростуть на тваринах і рослинах, як і на будь-якій іншій поверхні. Проте, їх ріст може бути збільшеним теплотою і потом, і значна кількість цих організмів на тілі людини — результат виділень тіла.
[ред.] Мутуалізм
Див. також: Нормальна флора
Певні бактерії формують тісні просторові асоціації, істотні для свого виживання. Один приклад такої мутуалістичної асоціації, назваий міжвидовим переносом водню, зустрічається між кластерами анаеробних бактерій, які споживають органічні кислоти і виробляють газоподібний водень, і метаногенними археями, які споживають водень [88]. Ці бактерії не в змозі споживати органічні кислоти і рости, коли водень накопичується в їх оточенні, і тільки у тісній асоціації з воднево-споживаючими археями, які утримують концентрацію водню достатньо низькою, ці бактерії можуть рости.
У ґрунті мікроорганізми ризосфери (зона, яка включає поверхню коренів і ґрунт, що торкається цього коріння) проводять фіксацію азоту, перетворюючи газоподібний азот на азотні сполуки [89]. Таким чином, бактерії забезпечують рослини зручною для поглинання формою азоту, яку рослини не можуть виробляти самостійно, отримуючи від рослин необхідні для життєдіяльності бактерій органічні сполуки. Інші бактерії знаходяться як симбіонти в тілі людини та інших тварин. Наприклад, в здоровій людині постійно присутні понад 1 тис. видів бактерій, більшість з яких, флора кишківника, можуть сприяти імунітету кишківника, синтезувати вітаміни (наприклад, фолієву кислоту, вітамін K і біотин) та розчиняти склідні важкорозчинні вуглеводи [90][91]. Інші бактерії, які пропонують вигоду людині-хазяїну, включають види Lactobacillus, які перетворюють молочні білки на молочну кислоту в кишківнику [92]. Присутність флори кишківнику також перешкоджає зростанню потенційно хвороботворних бактерій (зазвичай через конкурентне пригнічення останніх), і тому деякі з цих бактерії продаються як пробіотичні харчові добавки [93].
[ред.] Паразитизм
Див. також: Бактерії та здоров'я людини
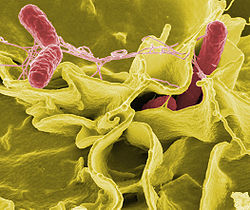
Електронна мікрофотографія з підсиленими кольорами, Salmonella typhimurium (червона) вторгується до людських клітин в культурі
Якщо бактерії формують паразитичну асоціацію з іншими організмами, вони класифікуються як патогени або хвороботворні бактерії. Хвороботворні бактерії є важливою причиною хвороб і смерті людей і викликають такі інфекції як правець, черевний тиф, дифтерит, сифіліс, холера, харчові отруєння, проказа, чума і туберкульоз. Бактерії часто стають причинами хвороб, які протягом довгого часу вважалися викликаними іншими причинами, наприклад, тільки на початку 1990-х років було відкрито, що причиною виразки шлунку є бактерія Helicobacter pylori. Бактеріальні хвороби також важливі в сільському господарстві: бактерії викликають багато хвороб рослин і тварин (хвороба Джонса, мастит, сальмонельоз, сибірська виразка).
Кожний від патогенних бактерій має характерну картину взаємодії з людиною-хазяїном. Деякі бактерії, наприклад, Staphylococcus або Streptococcus, можуть викликати інфекції шкіри, пневмонію, менінгіт і навіть сепсис, системну запалювальну відповідь, що приводить до шокового стану, масивної вазоділатації і смерті [94]. При тому, що ці організми — частина нормальної людської флори і зазвичай існують на шкірі або в дихальному тракті без породження будь-якої хвороби. Інші організми незмінно викликають хворобу в людині, наприклад Rickettsia, які є облігатними внутрішньоклітинними паразитами, здатними рости і відтворюватися тільки в межах клітин інших організмів. Один з видів Rickettsia викликає висипний тиф, інший — висипний тиф Скелястих Гір. Chlamydia, інший рід облігатних внутрішньоклітинних паразитів містить види, які можуть викликати пневмонію та інфекції сечового тракту і, можливо, коронарну хворобу серця [95]. Нарешті, деякі види, наприклад, Pseudomonas aeruginosa, Burkholderia cenocepacia, і Mycobacterium avium, є опортуністичними патогенами і викликають хворобу переважно в людей, що страждають пригніченням імунної системи, СНІД або кістозним фіброзом [96][97].
Бактеріальні інфекції часто можуть лікуватися за допомогою антибіотиків, які називаються бактерицидними, якщо вони вбивають бактерій, або бактеріостатичними, якщо вони тільки запобігають росту бактерій. Існує багато типів антибіотиків, і кожен клас інгібує процес, відмінний у патогені порівняно з хазяїном. Прикладом такої добірної токсичності є хлорамфенікол, який перешкоджує роботі бактеріальних рибосом, але не структурно відмінних рибосом еукаріотів [98]. Антибіотики використовуються як для лікування хвороб людей, так і в інтенсивному сільському господарстві, щоб просувати ріст тварин, але таке використання сприяє швидкому розвитку резистентності бактерій[99]. Інфекціям можна запобігти і антисептичними заходами, наприклад, стерилізуванням шкіри перед проникненням голкою шприца і правильним доглядом катетерів у медицині. Хірургічні і зубні інструменти також мають бути простерилізованими, щоб запобігти забрудненню і бактеріальним інфекціям. Дезинфікуючі засоби, наприклад, вибілювачі, використовуються для знищення бактерій та інших патогенів на поверхнях, щоб запобігти забрудненню і скоротити ризик інфекції.
[ред.] Економічне та екологічне значення
Див. також: Економічна важливість бактерій
Бактерії, зоктема Lactobacillus в комбінації з дріжджами і пліснявою, протягом тисяч років використовуються для виробництва продуктів бродіння, наприклад сиру, солених овочів, соєвого соусу, оцту, вина і кефіру [100][101].
Здатність бактерій руйнувати різноманітні органічні сполуки використовується в переробці відходів і біоремедіації. Бактерії, здатні до травлення вуглеводнів, використовуються для збирання розлитої нафти [102]. Наприклад, після катастрофи танкера Ексон Валдес в затоці Принца Вільгельма була розлита значна кількість нафти. В спробі очистити узбережжя від нафти на деякі пляжі були викинуті культури бактерій, які показали свою ефективнісь на не дуже сильно забруднених територіях. Бактерії також широко використовуються для біоремедіації промислових токсичних відходів [103]. У хімічній промисловості бактерії найбільш важливі у виробництві енантіомерно чистих хімічних речовин для використання у фармацевтиці або сільському господарстві [104].
Бактерії можуть також використовуватися замість пестицидів в біологійній боротьбі з шкідниками. Частіше за все для цього використовується Bacillus thuringiensis (також відома під назвою BT), грам-позитивна ґрунтова бактерія. Підвиди цієї бактерії використовуються як інсектицид, специфічний до лускокрилих, що продається зараз комерційно [105]. Через свою специфічність ці засоби вважаються екологічно дружніми, майже без негативного ефекту на людину, дику природу, запилювачів і на інших корисних комах [106][107].
Через їх здатність швидко рости і відносну легкість, з якою ними можна маніпулювати, бактерії широко використовуються у молекулярній біології, генетиці та біохімії. Створюючи мутації в бактеріальній ДНК і досліджуючи результуючий фенотип, учені можуть визначити функцію генів, ферментів і метаболічних шляхів у бактеріях, а потім застосовують це знання до складніших організмів [108]. Зараз розвинуті методи, що дозволяють накопичити значні набори даних про експресію багатьох генетичних продуктів одночасно та використовувати ці дані в математичних моделях біохімії цих організмів. Це досяжно відносно деяких найкраще вивчених бактерій, моделі бактерії Escherichia coli зараз активно досліджуються [109][110]. Розуміння бактеріального метаболізму і генетики дозволяє використання біотехнологій для виробництва за допомогою бактерій терапевтичних білків, наприклад, інсуліну, факторів росту або антитіл [111][112].
[ред.] Посилання