
Ім'я файлу: Характеристика мембранних ліпідів.docx
Розширення: docx
Розмір: 33кб.
Дата: 19.12.2022
скачати
Пов'язані файли:
ВІДКРИТИЙ МІЖНАРОДНИЙ УНІВЕРСИТЕТ РОЗВИТКУ ЛЮДИНИ.docx
Розширення: docx
Розмір: 33кб.
Дата: 19.12.2022
скачати
Пов'язані файли:
ВІДКРИТИЙ МІЖНАРОДНИЙ УНІВЕРСИТЕТ РОЗВИТКУ ЛЮДИНИ.docx
Характеристика мембранних ліпідів. Динаміка структурних елементів
мембрани. Білок-ліпідні взаємодії. Вода як складовий елемент
біомембран.
Ліпіди мембран представлені трьома основними класами полярних ліпідів: фосфолипидами
(Гліцеро- і сфінгофосфоліпіди), гліколіпідами і стероїдами.
Все мембранні ліпіди (незважаючи на відмінність у складі) є амфіфільних молекулами, побудовані за єдиним планом і мають дві області, що відрізняються спорідненістю до води: гідрофобні радикали (хвости) і полярні головки.
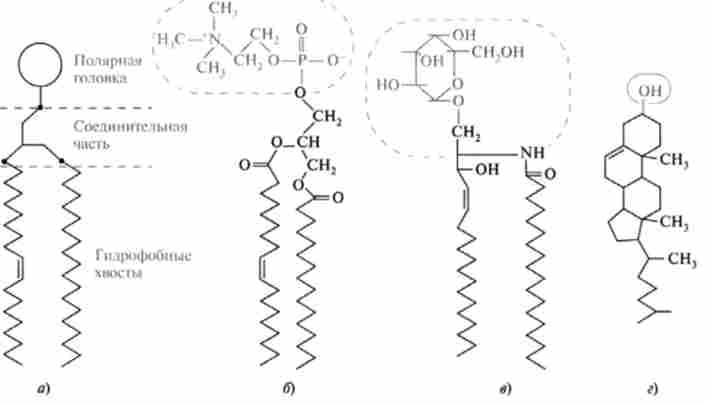
Схематичне зображення мембранних ліпідів (а) і формули фосфатидилхолина (б), галактоцераміда (в), холестеролу (г)
Подібні амфіфільних молекули у водному середовищі прагнуть до агрегації таким чином, що ліпофільні ділянки молекул (хвости), прагнучи потрапити в гідрофобну фазу, утворюють суцільні неполярні області, а полярні формують кордон між гідрофобною фазою і водою. Цей процес відбувається спонтанно, і серед асоціатів найбільш відомі мономолеку- лярні плівки (моношарів), міцели і бімолекулярні ліпідні шари.
Формування асоціата того чи іншого типу визначається перш за все співвідношенням розмірів полярної і неполярний частин молекули. Для фосфо і гликолипидов, які є основними компонентами біомембран, термодинамічно найбільш вигідно формування бислоя або бімолекулярного ліпідного шару.
Приблизний ліпідний склад різних клітинних мембран
Відсоток від загального вмісту ліпідів
| ліпіди | Плазмат. мембрана | мієлін | Зовнішня і внутр. мембрана мітохондрій | ЕПР | В. СОН | |||||
| клітин печінки | еритроцитів | |||||||||
| холестерол | 17 | 23 | 22 | 3 | 6 | 0 | ||||
| Фосфатиди мул етаноламін | 7 | 18 | 15 | 35 | 17 | 70 | ||||
| Фосфатіліл сери н | 4 | 7 | 9 | 2 | 5 | сліди | ||||
| Фосфат ід мул хол і н | 24 | 17 | 10 | 39 | 40 | 0 | ||||
| Сфінгоміслін | 19 | 18 | 8 | 0 | 5 | 0 | ||||
| гліколіпіди | 7 | 3 | 28 | сліди | сліди | 0 | ||||
| інші | 22 | 13 | 8 | 21 | 27 | 3 | ||||
Полярні групи ліпідних молекул утворюють дві гідрофільні поверхні, що відокремлюють внутрішню вуглеводневу фазу від водного середовища. Товщина ліпідного шару визначається перш за все довжиною вуглеводневих ланцюгів і зазвичай варіює в межах 4-5 нм. Присутність в вуглеводневих ланцюгах подвійних зв'язків в цис-конфігурації, бічних метильних груп і інших заступників порушує щільність упаковки молекул і призводить до зменшення товщини бислоя.
Головну частину ліпідної фракції складають фосфоліпіди (70-80%), крім них в мембрани входять гліколіпіди, холесте- рол, а також нейтральні ліпіди (моно-, ди-, триацилгліцеролів). Зміст останніх в плазматичній мембрані може становити до 16-20%; нейтральні ліпіди здатні збільшувати пружність і механічну міцність бішару. Плазматичну мембрану відрізняє також високий вміст холестеролу (15-20%), і лише дуже незначна його кількість знаходиться у внутрішньоклітинних мембранах (3-6%).
Білок-білкова взаємодія — зв'язування (зазвичай оборотне) білків, їх вплив один на одного, та дослідження цих процесів з точки зору біохімії, передачі сигналів і теорії керування.
Білок-білкова взаємодія важлива для багатьох біологічних процесів. Наприклад, сигнали від навколишнього середовища клітини передаються до її внутрішніх частин за допомогою білок-білкової взаємодії та взаємодії білків з сигнальними молекулами. Цей процес, що називається передачею сигналів, грає фундаментальну роль в багатьох біологічних процесах і в багатьох хворобах (наприклад, таких як рак).
Білки можуть взаємодіяти і протягом довгого часу, формуючи білкові комплекси та допомагаючи транспорту інших білків (наприклад при ядерному транспорті через ядерні пори від цитоплазма клітини до її ядра або навпаки), або можуть взаємодіяти протягом короткого часу, наприклад, змінюючи інші білки в результаті ферментативної реакції (наприклад, білкові кінази фосфорилюють певні інші білки). Ця модифікації білків можуть змінювати взаємодію з іншими білками. Наприклад, деякі білки з SH2-областями зв'язуються з іншими виключно у фосфорильованому стані.