
ОДЕСЬКИЙ НАЦІОНАЛЬНИЙ УНІВЕРСИТЕТ імені І. І. МЕЧНИКОВА
Біологічний факультет
Кафедра мікробіології
КУРСОВА РОБОТА
на тему: «Галофітні актинобактерії та їх чутливість до металів»
Використання галофільних актинобактерій у біоремедіації ґрунтів, забруднених важкими металами
Студентки ІІІ курсу
денного відділення
Кретової Катерини Олегівни
Керівник: канд. біол. наук, доцент
Зінченко Оксана Юріївна
Оцінка за шкалами:
національною _______
ЕСТS ______ Кількість балів:______
Члени комісії
Протокол засідання кафедри
№ ___ від “____” ______2022 р. _________ __________________
(підпис) (прізвище та ініціали)
_________ __________________
(підпис) (прізвище та ініціали)
_________ __________________
(підпис) (прізвище та ініціали)
Одеса – 2022
АНОТАЦІЯ
Курсову роботу «Використання галофільних актинобактерій у біоремедіації ґрунтів, забруднених важкими металами» викладено на 28 сторінках друкованого тексту, вона включає 1 малюнок. У роботі наведено посилання на 26 публікацій кирилицею та 8 латиницею.
ЗМІСТ
ОДЕСЬКИЙ НАЦІОНАЛЬНИЙ УНІВЕРСИТЕТ імені І. І. МЕЧНИКОВА 1
ВСТУП 4
РОЗДІЛ 1. ОГЛЯД ЛІТЕРАТУРИ 5
1.1.Актинобактерії та їх поширення в природі 6
1.2.Особливості систематики типу Actinobacteria 10
1.3.Галофільні актинобактерії 13
1.4. Вплив металів на галофітні актинобактерії 16
1.4.Механізми стійкості актинобактерій до впливу металів 20
УЗАГАЛЬНЕННЯ 23
СПИСОК ВИКОРИСТАНИХ ДЖЕРЕЛ 24
ВСТУП
Як і рослини, деякі види бактерій, зокрема актинобактерії пристосувалися до життя на засолених ґрунтах. Ця група мікроорганізмів має назву галофільних актинобактерій. Засолені ґрунти займають величезні території як у світі в цілому, так і в Україні. Щороку через неправильні сільськогосподарські процедури, особливо при ірраціональному зрошенні, їх територія розширюється. Саме тому важливо знати властивості таких ґрунтів, в тому числі склад та властивості угруповань мікроорганізмів у них, для вирішення питань, пов’язаних із засоленням та ефективним використанням. Традиційно вважалося, що міцеліальні актинобактерії не можуть займати природні екологічні ніші, що характеризуються екстремальними умовами, зокрема, надмірно засолені ґрунти, проте в подальших дослідженнях було встановлено, що це не так [4].
Метали, зокрема важкі метали, є небезпечними забруднювачами середовища. Ці речовини викликають зміну організації системи угрупувань бактерій та призводять до збіднення загального біорізноманіття видів, а також переважання видів, стійких до важких металів по відношенню до первісних спільнот у ґрунтах, не забруднених металами. Під впливом іонів металів в клітинах актинобактерій можна спостерігати різноманітні зміни, які в першу чергу пов’язані з їх морфологічними особливостями. Відомо, що навіть невеликі концентрації металів можуть прискорювати метаболізм, а також посилювати розвиток бактеріальних клітин, але великі дози одних і тих же металів надають токсичну дію на клітини [7-9].
Вплив металів на різні види мікроорганізмів різний та індивідуальний. У зв'язку з цим необхідно враховувати існуючі та небезпечні концентрації металів. Прояв токсичної дії важких металів на актинобактерії може бути різним, це залежить від фізико-хімічних властивостей металу, а також наявності інших чинників, які негативно вливають на мікроогранізм. Саме тому важливим є вивчення впливу металів, зокрема важкі метали на галофітні актинобактерії, адже вони зростають в засолених ґрунтах та зазнають подвійного впливу [2]. Визначення біологічних ефектів важких металів у клітинах галофільних актинобактерій стало підґрунтям для дослідження можливості їх використання як основи засобів для біоремедіації ґрунтів.
Метою роботи був аналіз особливостей пристосування актинобактерій до росту на засолених ґрунтах в присутності підвищених концентрацій важких металів та можливостей використання галофільних актинобактерій ля біоремедіації таких ґрунтів.
РОЗДІЛ 1. ОГЛЯД ЛІТЕРАТУРИ
Актинобактерії та їх поширення в природі
Актинобактерії – це група грампозитивних бактерій з високим вмістом гуаніну і цитозину. Здебільшого, вони є наземними або водними мешканцями. Актинобактерії є домінуючим типом бактерій [19].
Актинобактерії населяють ґрунти, а також прісні і морські води. Вони відіграють важливу роль у розкладанні органіки, такої як целюлоза і хітин, і таким чином беруть участь в циклі органіки і у вуглецевому циклі. Це поповнює запас поживних речовин в ґрунті, що важливо для формування перегною. Інші актинобактерії, в тому числі деякі патогенні, такі як мікобактерії, стрептоміцети, населяють рослини і тварин. Деякі актинобактерії утворюють розгалужені нитки, схожі на міцелій. Через це вони спочатку були класифіковані як актиноміцети. Більша кількість видів аеробні, але деякі, такі як Actinomyces israelii, можуть існувати в анаеробних умовах [13].
Окремі види актинобактерій відповідають за специфічний запах, що виходить з ґрунту після дощу. В основному це явище відбувається в районах з теплим кліматом. Актинобактерії відомі виробленням вторинних метаболітів, і тому є об'єктом вивчення у фармакології та промисловій мікробіології. У 1940 році Зелман Ваксман виявив, що ґрунтові бактерії, які він вивчав, виробляють антибіотик стрептоміцин, який став першим етіотропним препаратом для лікування туберкульозу, за що вчений отримав Нобелівську премію. З тих пір тисячі природних антибіотиків були виявлені серед метаболітів ґрунтових мікроорганізмів, особливо роду Streptomyces [25].
Метаболіти деяких видів актинобактерій (наприклад, Streptomyces griseus) зумовлюють запах ґрунту після оранки або дощової погоди, в основному в більш теплих кліматичних зонах. Багато дослідників (Красильников, 1950; Еріксон, 1953; Превот, 1953; і т.ін.) відзначили їх схожість з грибками і розглядали як проміжну форму між грибками і бактеріями [17, 28].
Вивчення ультраструктурної організації клітин актиноміцетів, а також аналіз хімічних компонентів дозволили віднести їх до прокаріотів і вважати їх однією з груп бактерій «в широкому сенсі».
Актиноміцети – одна із найбільших груп актинобактерій, їх клітини утворюють розгалужені вирости, а деякі мають міцелій товщиною 0,5-2 мкм, що зазвичай з'являється в екстремальних умовах. Клітинна мембрана схожа на мембрану грампозитивних бактерій. У клітині є нуклеоїд, вакуолі, включення, цитоплазматична мембрана утворює мезосоми [28-30].
При поверхневому культивуванні вони утворюють двофазний міцелій – в субстраті і на поверхні (останній диференційований). Актинобактерії розмножуються фрагментами міцелію або спорами. Спори утворюються безстатево на повітряному або субстратному міцелії і розташовуються поодинці, парами, ланцюгами або концентруються в спорангії. Деякі з них можуть бути мобільними [6].
Більшість актинобактерій є гетеротрофами і зазвичай невимогливі до джерел живлення. По відношенню до кисню вони в основному є аеробами, хоча серед патогенних зустрічаються і анаероби. Зазвичай вони розвиваються при нейтральній реакції середовища, але зустрічаються і ацидофільні форми (кислотостійкі) і лугостійкі, багато з них витримують висихання і засолення.
Актинобактерії широко розповсюджені в повітрі, водоймах, рослинних і тваринних залишках, ґрунтах, особливо безплідних – сухих, пустельних, сольових, кам'янистих, які знаходяться на перших стадіях ґрунтотвірного процесу. Вони беруть участь в перетворенні органічних і мінеральних речовин, також є продуцентами багатьох фізіологічно активних речовин: ферментів, амінокислот, вітамінів, антибіотиків, збагачують ґрунт азотом [8].
Актинобактерії дуже різноманітні за своєю морфологією та фізіологією. У цій групі зустрічаються наступні форми бактерій: коки; паличкоподібні; розгалужені нитки; з розвиненим міцелієм.
Усі актинобактерії грампозитивні та не утворюють ендоспор. Однак, наявність екзоспор – дуже поширена їх риса. Більшість актинобактерій є аеробами, проте серед них існують також факультативні та облігатні анаероби. Актинобактерії населяють різні середовища і відповідно мають різні типи живлення та обміну речовин. У цій представників цієї таксономічної групи зустрічаються всі типи живлення бактерій, окрім фототрофного [4-7].
Міцелій актиноміцетів утворений гіфами. Гіфи являють собою розгалужені тонкі нитки, які подовжуються в результаті верхівкового росту. Гіфи містять велику кількість нуклеоїдів.
Поділ клітин не може відбуватися в процесі розгалуження та росту гіф, але можуть утворюватися перегородки. Залежно від типу перегородок міцелій ділиться на 2 види:
без перегородок (несептований);
септований – містить перегородки, які здебільшого розташовуються в поперечному напрямку.
Міцелій може бути як стабільним, так і періодично фрагментованим на палички або коки, як в роду Nocardia. Розгалуження тонких ниток (гіф) в різних групах актиноміцетів виражене різною мірою. Актиноміцети здатні утворювати як субстратний, так і повітряний міцелій [25].
Актинобактерії – це бактерії з дуже високою біосинтетичною активністю. Вони здатні виробляти величезну кількість біологічно активних сполук, в тому числі і антибіотиків. З цієї причини мікробіологія актинобактерій займає особливе місце в біотехнології. Більшість природних антибіотиків були виділені з цієї групи мікроорганізмів.
Здатність утворювати тонкі нитки не є унікальною особливістю актиноміцетів, так як деякі представники інших груп мікроорганізмів мають подібну будову. Наприклад, рід Hifomicrobium, що відноситься до групи Proteobacteria [29-30].
Розмноження міцеліальних актиноміцетів може відбуватися трьома способами: фрагментація гіф, диференціація спор, Починаючим;
Спори – це утворення з потовщеною клітинною стінкою оточені гідрофобним покривом. Вони можуть бути як рухливими, так і нерухомими. Місцем мешкання більшості актинобактерій є ґрунт, а також прісноводні і морські мули. Зустрічаються представники, що живуть в екстремальних умовах з високим вмістом солі або високими температурами (галофіли, термофіли) [18].
Середовищем існування деяких актиноміцетів є тіло тварин і людини. Такі представники можуть бути патогенними для людини і тому є предметом вивчення медичної мікробіології. Актинобактерії можуть викликати такі захворювання: ендокардит, фарингіт, пародонтоз, легеневий нокардіоз, актиноміцетоз, туберкульоз, проказа.
Серед актинобактерій зустрічаються і збудники захворювань рослин, наприклад, рід Streptomyces, який викликає картопляну паршу. Деякі представники є симбіонтними бактеріями. В екологічній мікробіології актинобактерії вважаються найважливішими учасниками кругообігу речовин. Більшу частину цієї функції виконують саме актинобактерії, які живуть в ґрунті [21].
Отже, актинобактерії є типом грампозитивних бактерій, здебільшого вони займають водне та наземне середовище існування. Актинобактерії мають велике економічне значення для людей, оскільки сільське господарство і ліси залежать від їх внеску в ґрунт екосистеми, вони розкладають органічну матерію мертвих організмів. Деякі види живуть у симбіозі з рослинами, колонізуючи коріння та фіксуючи азот для рослин в обмін на доступ до деяких рослинних сахаридів. Інші представники є збудниками хвороб людини або, навпаки, джерелом отримання антибактеріальних, протигрибкових, противірусних засобів, імуномодуляторів, протипухлинних препаратів та інгібіторів ферментів. Продукти біосинтезу актиноміцетів знайшли своє застосування і в сільському господарстві у складі інсектицидів, гербіцидів, фунгіцидів та речовин, що стимулюють ріст рослин та тварин.
Особливості систематики типу Actinobacteria
З моменту свого відкриття у 70-тих роках XIX ст. і до середини 70-х років XX століття актиноміцети («променисті грибки») включали в основному міцеліальні грампозитивні бактерії, які мають зовнішню морфологічну схожість з мікроскопічними грибами, якими займався Красильніков. З ряду причин, включаючи різноманітність морфології, хімічний склад полімерів і молекул клітинної стінки, а також підвищений інтерес до цієї групи мікроорганізмів з боку біотехнології, систематика цієї групи вже в передмолекулярну епоху таксономії була однією з найбільш передових в порівнянні з більшістю інших груп прокаріотичних мікроорганізмів [19, 28].
Термін «актинобактерії» з'явився в систематиці на початку 80-х років XX століття, в початковий період переходу від класифікації, яка враховує в основному морфологічні та індивідуальні біохімічні особливості, до системи, заснованої на хемотаксономічних характеристиках і, особливо, даних про секвенування генів рРНК 16S [28].
Зокрема, за допомогою цих методів було встановлено, що ряд стрижневих і кокоїдних грампозитивних бактерій з високим вмістом Г + Ц – пар в ДНК (роди Arthrobacter, Micrococcus, Microbaclerium, Cellulomonas та ін.) є близькими родичами «класичних» міцеліальних актиноміцетів.
Такі бактерії в 80-х роках розглядалися як частина порядку Actinomycetales як колективна група супраномічного рівня, названа «актинобактеріями» – спочатку без надання цьому терміну формального таксономічного і номенклатурного статусу [26-28].
Особливо гостро виявилися складнощі систематики актиобактерій на рівні виду в роду Bаertus, який є найбільш численним за видовим складом. Ідентифікація геномоїдів, пошук надійних рис, що диференціюють геномогрупи на фенотиповому рівні, і подальший перегляд таксономічної структури роду – найактуальніші завдання систематики цих організмів в найближчому майбутньому.
Ідентифікація актинобактерій здійснюється по-різному. У класичній мікробіології для їх ізоляції і обліку використовується метод посіву з розведення ґрунтових суспензій та інших матеріалів на щільне живильне середовище. Туди, як правило, додаються речовини, які обмежують ріст грибків і неміцеліальних бактерій [15].
Для попередньої ідентифікації актиноміцетів використовується мікроскопія колоній на живильному середовищі, потім для отримання чистих культур відбираються окремі колонії. Ці культури повинні мати специфічні набори ознак, характерних для специфічних таксонів актинобактерій: культуральні (особливості росту на різних субстратах), фенотипові і генотипові [10-12].
Зазвичай враховуються такі фенотипові особливості:
морфологічні (наявність фрагментації міцелію, утворення спор на субстраті або повітряному міцелії, кількість спор в ланцюгах, наявність одиночних спор, спорангії)
хемотаксономічна (наявність L- або мезо-ізомеру діамінопімелінової кислоти (DAPk) і диференціювання цукрів в гідролізатах цілих клітин).
Сучасна систематика передбачає обов'язковий розгляд молекулярно-генетичних ознак – як мінімум, визначення послідовності гена 16S rRNA [30].
Фенотипово актинобактерії поділяють на 2 групи:
нижчі актиноміцети – не утворюють спор. До цієї групи входять поодинокі бактерії міцеліальної і неміцеліальної структури;
вищі актиноміцети – утворюють екзоспори і включають в себе тільки міцеліальні форми (як одноклітинні, так і багатоклітинні);
Виходячи з аналізу схожості гена 16-s-rRNA, 130 родів типу актинобактерій об'єднані в один клас – Actinobacteria, в якому виділяють 5 підкласів, 6 порядків і 10 підрядів.
Перші чотири підкласи не сильно розгалужуються і містять лише один рід кожен. П'ятий підклас, Actinobacteridae, ділиться на два порядки: Actinomycetales (актиноміцети); Bifidobacteriales (біфідобактерії).
У порядок Bifidobacteriales входить однойменна родина з 3 родами, а порядок Actinomycetales ділиться на 10 підрядів:
Actinomycinae.
Micrococcineae.
Corinebacterine.
Micromonosporineae.
Propionibacterineae.
Pseudonocardineae.
Streptomycineae.
Streptosporangineae.
Francineae.
Glycomycineae.
Багато з цих груп відповідають фенотиповій класифікації актинобактерій на основі сукупності морфологічних, культурних, фізіологічних і біохімічних ознак. Фенотипові групи актиноміцетів описані в другому томі визначника бактерій Берджі.
Нокардіподібні актиноміцети – сюди входять актинобактерії, міцелій яких періодично розщеплюється на палички або коки. Грампозитивні аероби не утворюють справжніх спор. Здатні утворювати конідії. Відсоток пар GC в цій групі варіюється від 63 до 79. Типовим представником є рідNocardia..
Актиноміцети з багатоклітинними спорангіями –. у таких мікроорганізмів поділ гіфів здійснюється в обидві сторони (поздовжній і поперечний), що призводить до утворення багатоклітинних спорангіїв. Два роди характеризуються відсутністю нитчастого міцелію [16-18].
Всі представники – хемоорганотрофи і мезофіли. Характерними місцями проживання є ґрунт, вода і шкіра ссавців.
Актинопланети пристосовані до життя у водному середовищі. Для них характерна наявність рухливої стадії в життєвому циклі. Міцелій утворюється септичний (в основному субстрат, рідше – повітря). Наявність спорангіїв характерна для всіх представників, крім роду Micromonospora. Місцем проживання є прісна вода, ґрунт і мертві органічні залишки [17].
Стрептоміцети – для них характерний стабільний і добре розвинений повітряний міцелій, який не піддається фрагментації. Рід Streptomyces включає приблизно 500 видів.
Мадуромицети – ця група характеризується утворенням розвиненого субстрату міцелію, але спори утворюються тільки на повітряних гіфах. Спори утворюють короткі ланцюги або зустрічаються в спорангії.
Термомоноспори – представники цієї групи актиноміцетів відрізняються здатністю рости при температурі від 40 до 48 ℃. Для них характерний повітряний міцелій з рухомими або нерухомими спорами. Типовим представником є рід Thermomonospora [18].
Отже, систематика актинобактерій вперше набула загальних обрисів на початку 80-х років XX століття, проте вона базувалася лише на морфологічних особливостях. З часом систематика змінилася і в основі поділу були покладені генетичні та біохімічні ознаки, з часом домінуючим став молекулярно-біологічний підхід. Найбільшою групою актинобактерій є актиноміцети, що характеризуються наявністю міцелію.
Галофільні актинобактерії
Природний та штучні гіперзасолені середовища з концентрацією солі вдвічі більше, ніж морська вода (наприклад, солоні озера і випарні ставки) зустрічаються на всіх континентах. Вони характеризуються великою різноманітністю мікробіоти аборигенних видів, пристосованих до життя при високих концентраціях солей, в яких є представники архей, бактерій і еукаріотів [13].
По відношенню до концентрації солі мікроорганізми поділяються на
наступні фізіологічні групи:
негалофіли, які вимагають менше 1% NaCl;
галофільні, які можуть рости при високих концентраціях солі, наприклад, 25% NaCl;
слабкі галофіли, найкращий ріст постерігається при 1-3% NaCl в середовищі;
помірні галофіли, оптимальний ріст яких знаходиться при вмісті в середовищі NaCl на рівні 3-15%;
екстремальні галофіли, для оптимального зростання яких потрібно 15-25% NaCl (Кушнер, Камекура,1988).
Досягнення в методології молекулярно-генетичних досліджень та біоінформатики значно розширили розуміння особливостей обміну речовин мікроорганізмів в надзвичайно солоних ґрунтах. Для ряду галофільних мікроорганізмів показано здатність до розкладання вуглеводнів та інших токсичних хімічних речовин, що дає можливість розглядати їх як перспективні засоби біоремедіації різних середовищ від складних забруднень [1-5].
Крім екологічного аспекту застосування екстремофілів, ці мікроорганізми користуються попитом в біотехнології. Наприклад, галофільні гамма-протеобактерії родин Halomonadaceae (включаючи членів родів Aidingimonas, Carnimonas, Chromohalobacter, Кобетія, Галомона, Галоталея, Кушнерія, Modicisalibacter, Salinicola і Zymobacter) є продуцентами солетолерантних ферментів (амілаза, протеаза і нуклеаза) [19].
Полісахариди бактеріальної поверхні мають великий потенціал для застосування в процесах видобутку нафти, фармацевтичній та харчовій промисловості. Деякі з них їх, зв'язуючись з токсичними сполуками, такими, як фенол, фосфорорганічні сполуки та інші, що утворюються при виробництві або використанні пестицидів і гербіцидів, сприяють їх подальшому розкладанню і, таким чином, очищенню від них навколишнього середовища.
Згідно з даними літератури [9], стрептоактиноміцети здатні розвивати високий осмотичний тиск в клітині, що пояснює їх значне поширення в засолених ґрунтах. У гумусово-накопичувальних солончакових горах гідроморфного солончака кількість актиноміцетів склала сотні тисяч по відношенню до 1 г ґрунту, і їх кількість відповідала 29% від загальної кількості бактерій.
Дослідження показують, що спостерігається зменшення кількості бактерій по профілю солончака, що також характерно для зональних ґрунтів – чорноземів [16]. Частка актиноміцетів в прокаріотичному мікробному співтоваристві перехідного гумус-накопичувального генетичного горизонту солончака залишається досить високою (40%). Отримані результати узгоджуються з інформацією про те, що актиноміцети складають еколого-трофічні групи мікроорганізмів, найбільш широко представлених у солончаках.
Вниз по ґрунтовому профілю соляних поверхонь загальна кількість бактерій і кількість актиноміцецетів значно зменшується тільки в горах. Так, дослідники показують, що частка міцеліальних актинобактерій у прокаріотичному мікробному співтоваристві досліджуваних засолених ґрунтів, розташованих на посушливих територіях України, досить значна, досягає 40% і відповідає тій, що у чорнозему.
Що стосується видового різноманіття представників роду Streptomyces то вони представлені в невеликих кількостях на території солончаків. Відзначається наявність видів, об'єднаних в 5 секцій і рядів – секція Cinereus серії Aureus (стрептоміцети видів, що належать до цього розділу і ряду, складають 37% від загального комплексу актиноміцетів ґрунту); секції серії Cinereus Chromogenes (27% комплексу); Альбусні секції серії Альбоколят (18%); Альбусні секції серії Альбус (9%); секції серії Roseus Roseoviolaceus (9%). Поширеними видами в солончаках є Streptomyces violaceomacula-tus (секція Roseus серії Roseoviolaceus)[15].
У результаті мікробіологічних аналізів було виявлено, що в ґрунтах солончаків з високою травою кількість актиноміцетів переважає вміст грибів у 1,6 рази, а на середньотрав’ї і темно-каштанових злегка підсолених ґрунтах – актиноміцети переважають над грибами в 3,2 рази [9].
У численних дослідженнях ґрунтів показано, що максимальна кількість олігокарбофільних актиноміцет спостерігається в середині вегетаційного періоду рослин. Олігонітрофільні актиноміцети на солончаках мають максимальну бімасу також в середині вегетаційного періоду рослин, тільки на темно-каштанових ґрунтах з низьким вмістом солей максимальна кількість цих актиноміцетів спостерігається наприкінці літа. Найбільша кількість оліготрофних актиноміцетів на солончаках і темно-каштанових низькосолоних ґрунтах відзначається в середині літа [6-9].
Отже, кількість мікроорганізмів, зокрема галофільних актинобактерій, коливається залежно від ступеня засолення солончаків і наближених до солончакових ґрунтів, а також від флористичного складу рослинності. Велика кількість актиноміцетів зустрічається на темно-каштанових низькосолоних ґрунтах, за винятком олігонітрофільних актиноміцет, що пов'язано з низьким вмістом у ґрунті водорозчинних солі і збільшенням гумусових речовин.
1.4. Вплив металів на галофітні актинобактерії
Підвищений вміст важких металів в засолених ґрунтах призводить до зниження активності їх мікробних угруповань, родючості і, в кінцевому рахунку до зниження врожайності сільськогосподарської продукції культур. Експерименти показали, що кількість бактерій в кореневищі трави Діплахне фуска, вирощеної на ґрунтах з високим вмістом важких металів склав 1,0×107 КУО/г, що на кілька порядків менше, ніж у незабруднених ґрунтах [21].
У природних водоймах солі металів перебувають в розчиненому та адсорбованому станах, який визначається хімічним складом, температурою та рН води. Після потрапляння в воду солі нагромаджуються у вигляді карбонатів, сульфідів, фосфатів. Концентрація цих сполук у водоймі є досить різноманітною.
Мікробіологічні дослідження показують, що у ґрунтах, забруднених важкими металами, значно зменшуються біорізноманіття та біомаса мікробних спільнот. Як довгострокові, так і короткострокові забруднення негативно впливають на фізіологічну та біохімічну активність мікробіоти ґрунту, особливо дихання. У той же час деякі ризобактерії здатні виробляти ряд речовин, таких як антибіотики (в тому числі, фунгіциди), леткі ціаніди, фітогормони (МАК), сидерофори, що підвищують доступність важких металів для рослин і сприяють їх всмоктуванню корінням [24].
Вміст в середовищі важких металiв може спричиняти структурну перебудову мiкробоценозів ґрунтів. У них поступово зменшується кількість чутливих видів і залишаються ті, що здатні пристосуватися до високих концентрацій важких металів та інших шкідливих речовин. Іони важких металів після потрапляння через ґрунт до організму рослин через харчові ланцюги здатні потрапити в організм людини та тварин. Для зменшення забрудненості ґрунтів застосовують мелiоранти, здатні зв’язувати солі важких металів, перетворюючи їх в інші сполуки.
Одним з альтернативних методів очищення ґрунтів є біологічне лікування або біоремедіація, суть якої полягає в полягає у використанні для видалення забруднюючих речовин мікроорганізмів або інших біологічних систем. Дослідження показують, що ризобактерії можуть бути стійкимм до важких металів і грати значну роль у їх мобілізації або знерухомленні.
Встановлено, що вирощування актинобактерій в середовищах, забруднених важкими металами, призводить до зміни (зменшення або збільшення) розмірів бактеріальних клітин. Так, при вивченні морфометричних параметрів R. полностью ruber IEGM 70T в умовах забруднення середовища важкими металами встановлено збільшення довжини бактеріальні клітини (до 10 мкм) за впливу іонів кадмію і хрому, тоді як забруднення іонами
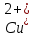 ,
, 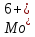
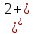 ,
, 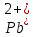
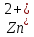 призводило до зменшення довжини клітин до 2 мкм [27].
призводило до зменшення довжини клітин до 2 мкм [27].Оцінюючи вплив токсичності навколишнього середовища, забрудненого важкими металами, та засоленість ґрунтів на життєздатність актинобактерій, можна виділити наступні закономірності: 1) штами бактерій по-різному реагують на токсичність важких металів; 2) життєздатність одних мікроорганізмів вище на носіях, забруднених свинцем і цинком, інших – міддю і кадмієм; 3) летальна концентрація поживних речовин при забрудненні свинцем, міддю і кадмієм встановлюється на рівні 0,3% і вище; 4) цинк має найменший інгібіторний вплив на виживання симбіонтів: при вирощуванні в середовищі з 0,01-0,1% вмістом цинку відзначається стимуляція росту, подальше збільшення концентрації цинку призводить до помітного гальмування життєздатності досліджуваних штамів [23-25].
Осадження сполук металів відбувається за допомогою мікроорганізмів при взаємодії з біогенним сірководнем, він утворюється при розкладанні органічних сполук, які містять у складі сірку у максимально відновленому стані. Метали також можуть зникнути при взаємодії з білками, тоді вони утворюють нерозчинні комплекси. Цей тип взаємодії найбільш характерний для іонів купруму. Внаслідок реакцій утворюється вуглекислий газ, це призводить до утворення нерозчинних сполук кадмію та плюмбуму (CdCO3, PbCO3) [19-21].
Плюмбум та багато інших металів здатні утворювати нерозчинні сполуки з бурштиновою кислотою. Акумуляція металів здійснюється як при високих, так і при незначних концентраціях металів. Культура Acidithiobacillus ferrooxidans (попередня назва Thiobacillus ferrooxidans) може рости й акумулювати метали при концентрації, що не перебільшує 30-40 г/л.
Акумуляція металів мікроорганізмами може здійснюватися й за рахунок зв’язування з клітинною стінкою. Метали здатні до нагромадження всередині клітин. У літературі є дані про внутрішньоклітинне нагромадження аргентуму, цинку, кобальту, кадмію. Таким чином, нагромадження металів в клітинах пов’язане з транспортними системами, причому в поглинанні металу, здебільшого, беруть участь елементи як пасивного так і активного транспорту [13-15].
Після того, як метали потрапили в клітину, вони зв’язуються з білками цитоплазми та внутрішніми структурами або утворюють нерозчинні продукти всередині клітини, включення. До прикладу, кадмій та ферум всередині клітин входять до складу сульфідів. Леткі сполуки металів утворюються тільки при взаємодії мікроорганізмів з окисненими сполуками меркурію, вони утворюються у вигляді металічного меркурію.
Метали здійснюють вплив на мікроорганізми на двох рівнях. Перший з них пов'язаний з нестачею металів, що необхідні для нормального функціонування бактерії. Другий пов’язаний із надлишком та шкідливою дією металів, а саме пригніченням діяльності ферментів. Причини інактивації ферментів можуть бути різними. Метали здатні зв’язуватися з активним центром та займати місце необхідних металів, тим самим руйнуючи білкову структуру.
У малих концентраціях солі металів є необхідними для актинобактерій, адже вони входять до складу ферментних систем. Вперше згубна дія металів на організм бактерій була описана в 1893 році, це явище отримало назву олігодинамічної дії та пояснювалося здатністю деяких металів у невеликих концентраціях викликати морфологічні зміни, а потім при збільшенні концентрації і загибель клітин мікроорганізмів. У концентраціях більше 1% метали викликають денатурацію білків і нуклеїнових кислот. У дуже високих концентраціях дія металів може бути специфічною, оскільки виявляється певна спрямованість у сприйнятті бактеріями окремих видів металів [5].
Отже, найбільше впливу металів піддаються процеси клітинного поділу, транспорт речовин та проникність мембрани. Аргентум і купрум здатні також чинити вплив на клітинну стінку, збільшуючи її проникність. Порушення цих процесів може призводити до порушення функцій клітини загалом, а в окремих випадках й до загибелі. Серед біохімічних процесів найбільший вплив виявлено щодо бродіння. Причиною загибелі клітин у таких випадках може бути повне пригнічення енергетичних процесів.
Механізми стійкості актинобактерій до впливу металів
Як відомо, повністю відмовитися від важких металів зараз практично неможливо, тому їх продовжують комплексно використовувати в багатьох галузях промисловості, що робить їх глобальними забруднювачами планети. У зв'язку з цим вміст важких металів в компонентах навколишнього середовища строго регламентується. Як відомо, важкі метали згубно впливають на мікроорганізми, а їх токсичність залежить від біодоступності важкого металу і поглинених доз [2].
Результати багатьох досліджень свідчать про високий рівень резистентності мікроорганізмів, ізольованих із хімічно забруднених ґрунтів навколо підприємств, до іонів важких металів. Ріст та розвиток досліджуваних ізолятів показує, що вони адаптуються по-різному до різних концентрації іонів міді і кобальту в живильному середовищі. Тобто було встановлено, що деякі з вибраних ізолятів стійкі до різних концентрацій іонів міді, інші – іонів кобальту.
Мікробіологічні процеси в ґрунті можуть призводити або до переведення сполук металів у розчинну форму (тобто підвищення їх біодоступності та потенційної токсичності) або до їх знерухомлення (тобто зменшення біодоступності). Ці процеси біотрансформації є важливими компонентами біогеохімічних циклів використовуваних металів.
Для біоремедіації зразків, забруднених важкими металами, використовують біоакумуляцію, що, як відомо, включає поглинання забруднювачів живою бактеріальною біомасою, грибів і мікроводоростей. Серед мікроорганізмів наявні форми, що здатні чинити опір дії деяких токсичних речовин. Окремі види можуть використовувати сполуки для своєї користі. Гени, що визначають стійкість до важких металів, часто знаходяться в плазмідах, іноді в хромосомах.
Вважають, що за стійкість до дії токсичних речовин в клітинах найчастіше відповідають плазміди [1-3].
Трансмісивні плазміди актинобактерій містять гени множинної стійкості – R-фактори, що спричиняють резистентність мікроорганізмів до кількох груп речовин – солей важких металів, а також антибіотиків, лікарських речовин та ін.
Такі плазміди відносяться до катаболічних. Катаболічні плазміди несуть гени синтезу ферментів, що деградують різні сполуки, наприклад, антибіотики або ксенобіотики (пестициди). Наприклад, деякі штами S. aureus в плазмідах містять гени стійкості до іонів кадмію. Часто, хоч і не завжди, вони пов'язані з генами стійкості до антибіотиків [23].
Ці гени кодують синтез ферментів – відповідних редуктаз. Можна виділити адаптивні індуцибельні редуктази, які утворюються при культивуванні штамів на середовищах з важкими металами, та конститутивні редуктази, які постійно продукуються мікроорганізмом, навіть при вирощуванні на звичайних середовищах.
Вплив факторів середовища призводить до інсерцій, дилецій або перерозподілу послідовності ДНК у плазмідах. Такі рекомбінаційні події змінюють структурну цілісність плазміди, і цим можуть впливати на експресію її генів шляхом їх переорієнтації щодо промотору або видалення інсерційно інактивованих послідовностей [30].
Рекомбінація може збільшувати метаболічні можливості мікроорганізму. Пластичність катаболічних плазмід забезпечує механізм, за допомогою якого обмін генетичним матеріалом може призвести до створення організму, здатного до ефективної утилізації нового субстрату у фазі експоненційного росту, тобто. у накопичувальній культурі. У природних спільнотах мікроорганізми, які спочатку не мали здатності утилізувати цю сполуку, у відповідь на присутність нового субстрату «реконструюються» шляхом перенесення генетичної інформації, що призводить до здатності утилізувати новий субстрат [4].
Антимікробний ефект важких металів заснований на здатності блокувати білки та інші органічні сполуки. Як антисептики у медичній практиці широко використовуються нітрат срібла (ляпіс), сульфат міді (мідний купорос) та хромат ртуті. Не рекомендується застосовувати для дезінфекції сполуки свинцю, миш'яку та ртуті, оскільки вони здатні акумулюватися в організмі людини.
Актинобактерії концентрують важкі метали на своїй поверхні та інших структурах. Концентрація важких металів всередині клітини актинобактерій може значно перевищувати концентрацію зовні. В одних випадках накопичення відповідних сполук виявляється летальним, а в інших – ні. На поглинання іонів металів можуть впливати фізіологічний стан клітин та умови довкілля [15-19].
У ряді випадків у мікроорганізмів виробилися способи видалення цих речовин із середовища шляхом їхнього осадження, адсорбції або випаровування. Такі реакції бактерій мають вагомий вплив на середовище, зокрема на його очищення, що є корисним для організмів, які не здатні боротися з токсичними речовинами самостійно.
Отже, галофільні актинобактерії займають особливе місце серед мікроорганізмів, які здатні до існування в екстремальних умовах, адже вони не тільки пристосовуються до умов росту на засолених ґрунтах, а й до дії металів ззовні. З чисельних праць випливає, що актинобактерії здатні перетворювати шкідливі речовини в інші види сполук, що не завдають такого важкого впливу на живі організми. Вони підтримують сталий розвиток в біоценозі, знижуючи токсичність речовин.
УЗАГАЛЬНЕННЯ
Актинобактерії є типом грампозитивних бактерій, що здебільшого займають водне та наземне середовище існування. Найбільшою групою актинобактерій є актиноміцети, що характеризуються наявністю міцелію.
Актинобактерії мають велике економічне значення для людей, оскільки сільське господарство і ліси залежать від їх внеску в розкладання органічної матерії мертвих організмів у ґрунтових екосистемах та у кругообігу хімічних елементів. Деякі види живуть у симбіозі з рослинами, а саме, в ризосфері та відіграють роль у підвищенні доступності азоту для рослин в обмін на доступ до деяких рослинних сахаридів. Окремі представники є збудниками хвороб людини, інші – джерелом отримання антибактеріальних, протигрибкових, противірусних засобів, імуномодуляторів, протипухлинних препаратів та інгібіторів ферментів. Препарати, до складу яких входять актинобактерії, використовуються в сільському господарстві як інсектициди, гербіциди, фунгіциди та речовини, що стимулюють ріст рослин та тварин.
Актинобактерії пристосовані до існування у різних умовах, зокрема, у ґрунтах з високим вмістом солей, а також забруднених хімічними полютантами. Ця особливість актинобактерій може бути використана для біоремедіації ґрунтів, забруднених, зокрема важкими металами. Важкі метали, на жаль, залишаються необхідними компонентами багатьох виробництв і є одними з найбільш значущих полютантів у глобальних масштабах.
Галофільні актинобактерії здатні не лише присостовуватися до існування у забруднених і засолених ґрунтах, а й перетворювати сполуки важких металів у низько- або нетоксичні.
СПИСОК ВИКОРИСТАНИХ ДЖЕРЕЛ
Агре Н.С. 1986. Систематика термофильных актиномицетов / –Пущино: 132 с.
Агре Н.С. и др. - 1972. Электронно-микроскопическое изучение спорообразования Micropolyspora viridinigra и Micropolyspora rubrobrunnea // Микробиология. - № 41. – С.854-858.
Артамонова В. С. и др. – 2010 Техногенное засоление почв и их микробиологическая активность // Сибирский экологический журнал.– №3. – С. 461–470.
Багаева Т. В. и др. 2005 Микробиологическая ремедиация природных систем от тяжелых металлов / – Казань: Казан. ун-т., – 56 с.
Балц Р. Х. 2005. Відкриття антибіотиків від актиноміцетів: чи відбудеться ренесанс за занепадом і падінням? / № 55. С.186 –196.
Бальц Р.Х. 2007. Протимікробні засоби з актиноміцетів // Назад у майбутнє.
Богачева А.С. 2011. Чувствительность цианобактерий к токсическому действию солей тяжелых металов / - Санкт – Петербург – 139 с.
Гришко В. Н. и др. – 2010. Структурно функциональные особенности сообщества актиномицетов в некоторых черноземах и засоленных почвах Украины // Почвоведение.– №2. – С. 221–228.
В.Н. Гришко, и др. – 2015 Мицелиальные актинобактерии засоленных почв аридных территорий Украины и России // Почвоведение. – № 1. – С. 81-86.
Добровольская Т.Г. 2002. Структура бактериальных сообществ почв / - Москва : ИКЦ "Академкнига", - 282 с.
Звягинцев Д.Г. 1980. Методы почвенной микробиологии и биохимии / - Москва : МГУ, - 224 с.
Звягинцев, Д. Г. и др. – 2008 Мицелиальные бактерии засоленных почв // Почвоведение.– № 10. – С. 1250-1257
Ибрахим И. М. и др. - 2018. Характеристика гликополимеров поверхности галофильных грамотрицательных бактерий Chromohalobacter salexigens EG1QL3 и Halomonas ventosae RU5S2EL // Известия Саратовского университета. Новая серия. Серия Химия. Биология. Экология. - Т. 18, - № 3. - С. 312–317.
Іутинська Г. О. та ін. -1999. Вплив міді на мікрофлору темно-сірого опідзоленого ґрунту і розробка способів його оздоровлення // Бюлетень сільськогосподарської мікробіології. - № 4. - С. 11–14.
Іутинська Г. О. та ін. - 2000. Відновлення біологічних функцій ґрунтів, забруднених важкими металами // Агрохімія і ґрунтознавство. С. 93–95.
Іутинська Г. О. та ін Токсичність і мутагенність важких металів - забруднювачів ґрунту // Сучасні проблеми токсикології. - № 2. - С. 53–56.
Квасніков Е. І. та ін - 1993. Біологія бактерій, яких використовують при очистці промислових стічних вод від важких металів // Мікробіологія. - Т. 55. - № 6. - С. 23–30.
Костина Л.В. - 2010. Аккумуляция солей тяжелых металлов клетками актинобактерий и использование Rhodococcus-биосурфактантов для мобилизации и извлечения тяжелых металлов из нефтезагрязненной почвы / Пермь : Микробиология, – 264 с.
Красильннков Н.А. – 1960 О принципах классификации актиномицетов. Биология отдельных групп актиномицетов - продуцентов антибиотиков // Труды инта микробиологии АН СССР. - № 8. - С.7-20.
Кузнецов В.Д. - 1973. Гомологические ряды наследственной изменчивости актиномицетов // Антибиотики. - № 7. – С.579-586.
Першина Е. В. и д.р. – 2012. Изучение структуры микробного сообщества засоленных почв с использованием высокопроизводительного секвенирования // Экологическая генетика. – Т.10. – № 2. – С. 32–39.
Петруша З. В. и д.р. – 1999. Біоіндикація забруднених важкими металами ґрунтів та заходи їх оздоровлення // Екологічність продукції АПК: економіка та технологія. - С. 206–209.
Сенцова О. Ю. и др. - 1985. Действие тяжелых металлов на микроорганизмы // Успехи микробиологии. - № 20. - С. 227–252.
Синдирева А.В. и др. - 2018. Экологическая оценка действия свинца в системе «почва – растение – животное» и разработка научно обоснованных приемов его детоксикации // Вестник КрасГАУ. - № 6. - С. 244–249.
Таширев А. Б. и др. - 1997. Аккумуляция тяжелых металлов микробным сорбентом // Мікробіологія. - Т. 59. - № 3. - С. 70 –79.
Таширев О. Б. - 2004. Взаємодія мікроорганізмів зі сполуками міді // Агроекологічний журнал. - № 1. - С. 42–46.
Babich H. et al - 1978. Toxicity of zinc to fungi, bacteria, and coliphages: influence of chloride ions // Appl. Environ. Microbiol. - V.36. P. 906–914.
Goodfellow M. et al - 1977. The actinomycetc genus RHodococcus a home for the 'rhodochrous' complex // J Gen Microbiol. P. 99-122.
Kushner D.J. et al – 1988. Physiology of halophilic eubacteria // Halophilic Bacteria. – V. 1. – P. 87-103.
M. Ventura et al – 2007. „Genomics of Actinobacteria: tracing the evolutionary history of an ancient phylum“// Microbiol. Mol. Biol. - T. 71. - V.3. P. 495–548.